

Does sleep prevent or promote forgetting?
01 - 10 - 2012
Abstract
A wide range of evidence supports the idea that sleep could have a role in memory consolidation. However, we are now gathering evidence that sleep could also have an active role in forgetting. The resulting model assumes that slow-wave sleep may globally weaken memory, but nevertheless strengthen relevant information over other. Thus, sleep might enhance competitive memories forgetting, a commonplace process that has already been described in human during wakefulness. Therefore, to promote memory consolidation, sleep may promote forgetting. However, recent behavioral studies fail to give consistent results for or against this model.

Table of Contents
Introduction
The idea that sleep improves memory processing has a long history and has been highly challenged and enriched these last decades. While Crick & Mitchison [1] early suggested that sleep could promote forgetting of daily unwanted memories, their hypothesis failed to reach researchers mind as most of the investigations reported opposite results, that is, sleep prevented forgetting. Now a wide range of evidence supports the sleep-related memory consolidation theory and coherent models considering the structural complexity of both memory and sleep begin to emerge [2,3]. Although current theory may acknowledge that passively forgetting some information would be a consequence of overnight consolidation of others memories [3], one might deplore that the relationship between sleep and forgetting has not been further investigated. Fortunately, the cognitive processes underlying forgetting are more and more understood [4], but it seems that research on forgetting and the one on sleep and memory do not benefit from each other yet.
This short note aims to review evidence for such a connection and to examine whether sleep could be related to an active forgetting process and also be compatible with the current sleep-related memory consolidation theory. Therefore, we will first analyze a part of the large amount of data on sleep, memory (especially the hippocampal-dependent declarative memory) and forgetting. We will then suggest a model, according to which sleep may promote a forgetting mechanism based on the selective retrieval of some memories over others. Finally, we will challenge this model with original studies published these past two years that tackled with the issue of sleep and forgetting.
Clues for a theoretical sleep-related forgetting
Synaptic plasticity and sleep stages
As hippocampal synaptic plasticity is very likely to be a major player of both memorization and forgetting mechanisms [5], a wide range of studies have investigated how it was modulated through the different states of vigilance. Long-term synaptic plasticity relies to a large extent on two well studied opposite mechanisms: long-term synaptic potentiation (LTP) and long-term synaptic depression (LTD) (see [6] for a review of the processes described below). It has been established that simultaneous activation of both pre-synaptic and post-synaptic membrane potential (i.e. as a consequence of the simultaneous firing of interconnected neurons) is able to strengthen synaptic efficacy. This potentiation is known to involve increase in Ca2+ concentration in post-synaptic compartment and the expression of the glutamate AMPA receptors. Conversely, the removal of the AMPA receptors underlies synaptic depression.
Data from many different fields such as biochemistry, genetics, electrophysiology have been collected and revealed a substantial effect of sleep on synaptic plasticity. First, it has been shown that synaptic potentiation was globally inhibited during slow-wave sleep (SWS, the early sleep stage which is characterized by synchronous neural activity) [7]. More precisely, the induction of new LTP is impaired but LTP which was previously triggered (i.e. concerning awake experiences) may be further processed [8]. This could be related to the low activity of all the main neural systems during SWS, especially the cholinergic system [9] and to the decrease in the expression of synaptic potentiation markers and the removal of AMPA receptors from the post-synaptic membrane [10]. Conversely, SWS might favor synaptic depression. This idea is more specifically supported by the similarity between the electrical activity which characterizes SWS (i.e. low frequency oscillations) and the one which artificially induce LTD (low frequency stimulation) [11]. Actually, recent studies managed to find correlation between slow wave activity (SWA) power and synaptic depression. For instance Lanté et al. provided evidences that SWA might directly lead to the removal of Ca2+-permeable AMPA receptors from synaptic membrane and the induction of LTD [12]. These electrophysiological data are correlated with the increase in expression during SWS of synaptic depression markers such as calcineurin, a phosphatase that especially targets AMPA receptors to promote their removal [13].
On the other hand, rapid eye movement sleep (REM sleep, the other main sleep stage which is characterized by a wake-like neural activity and muscle atonia) appears to promote potentiation of synaptic connections at the opposite of SWS-related synaptic plasticity. High cholinergic activity as well as increasing expression of immediate early genes involved in neuronal activation and synaptic plasticity are thus observed during REM sleep strongly suggesting an active role of REM sleep in LTP maintenance and induction [9,14]
To sum up, sleep synaptic plasticity greatly depends on the state of vigilance. SWS might globally promote synaptic depression and reduce LTP induction whereas REM sleep would favor some synaptic consolidation mechanisms. These results constitute a cornerstone of the sleep-related consolidation theory. Preventing new input to occur during SWS may thus minimize the likelihood that new information interfere with previously learned material (i.e. retroactive interference) and finally weaken previously encoded memories (see [15] for a detailed review). Therefore, SWS may protect memories from retroactive interference resulting in overnight better retrieval of these memories. However, we do not aim to study the consolidation processes further but rather to focus on how forgetting could occur in that context. Several works and in particular recent studies from Malleret et al. in rodent suggest that LTD is closely related to forgetting. Thus they showed on the one hand that genetic LTD impairment led to a damaged behavioral flexibility preventing the animal to forget previous experiences [16]. On the other hand, LTD enhancement seemed to reduce interference and facilitate the acquisition of new information [17]. Given that, it is likely that SWS-related LTD might promote forgetting of interfering memories. Such a hypothesis may be linked to the SWS-related synaptic downscaling proposed by Tononi & Cirelli [18] that we will now further analyze.
The synaptic homeostasis theory: a theory of forgetting?
It has been demonstrated in many species that the more one spends time awake, the more synaptic connections number and strength increase [10,19,20]. This increase in synaptic potentiation would be unsustainable on the long run because of the increasing demand in energy and space it implies. The synaptic homeostasis theory assumes that the synaptic overloading seen during wake is counterbalanced by a global downscaling occurring during SWS [18]. Among the arguments supporting this theory is the decrease in SWA power as the night goes on [21]. This decrease would reflect the fact that neurons are less likely to fire synchronously and suggest the global weakening in network associative strength. In addition, there is a positive correlation between the cerebral activity occurring during wakefulness and subsequent SWA power in SWS [22]. That is, the higher is the demand placed on the brain through the day, the more the subsequent sleep is refreshing. Moreover, such a renormalization of neural networks during SWS appears to be essential to restore cognitive abilities [23].
The model predicts for one neuron a proportional strength reduction of all its afferent synapses (i.e. synaptic downscaling). Thus, it would have three main consequences: (a) the global depression of the network, (b) the wipe out of the weakest connections, (c) the increase of the signal-to-noise ratio. However, it is to note that synaptic downscaling differs from the previously described LTD. While the later is synapse specific and could occur independently from prior synapse activity, synaptic downscaling involves most, if not all, synaptic connections of a single neuron and rely on their previous potentiation. Nevertheless, the underlying mechanisms of synaptic downscaling and LTD may share similarities [18].
As far memory processing is concerned, the synaptic homeostasis hypothesis would suggest that forgetting of irrelevant information would be linked to the consolidation of memories which were previously potentiated. In other words, competitive memories (interference) would be weakened resulting in a better retrieval of core (relevant) memories due to a higher signal-to-noise ratio. This hypothesis has been supported by behavioral study that showed lower effect of retroactive interference on word-pairs learning if the task was followed by sleep [24]. However, the same study demonstrated that sleep-related memory consolidation could also be stronger on poor encoded memories. That is, if a global forgetting effect of an eventual synaptic downscaling would occur, it might be impaired by local active consolidation processes. Therefore, there might be mechanisms that support this selective memory strengthening during SWS despite this sleep stage may promote global synaptic weakening. We will now discuss the hypothesis that some memories are more likely to be remembered than others through the analysis of recent works on neural networks level.
Are memories tagged for subsequent sleep processing?
In order to inquire whether sleep could selectively promote the consolidation of some memories over others, we will analyze the findings of electrophysiological approaches of (a) daily forgetting mechanisms and (b) selective sleep processes which might be involved in memory strengthening. This is an important point of our work to the extent that these two research fields are scarcely tied together when one offers theoretical basis of sleep memory function.
On the one hand, the works of A.D. Wagner and his colleagues have brought fundamental results concerning the knowledge of forgetting. Actually, they suggest that several distinct forms of forgetting exist relying on different mechanisms [4]. One of them is the forgetting of competitive memories in order to overcome interference. That is, irrelevant memories may be selected against and then forgotten as one achieves the retrieval of the desired information. Wagner and colleagues describe this kind of forgetting as follows: forgetting by resolving competition. This selective process is thought to involve the activation of the prefrontal cortex which would discriminate the information to be remembered from the one to be forgotten [25]. Moreover, hippocampal neural networks that were activated during an episodic memory encoding are likely to be reactivated when associated competitors are acquired and thus prevent the forgetting of the original information [26]. We previously have suggested that sleep may promote kind of forgetting. Now we assume that forgetting by resolving competition is the process that would most likely support this hypothetical sleep-related forgetting. The underlying mechanisms that would drive discrimination between relevant and competitive information during sleep may thus share similarities. On the other hand, it has been investigated whether similar selection mechanisms are tied to sleep- related memory consolidation. Many studies showed that neurons that have been related to the encoding of some memories tend to be reactivated during subsequent SWS [27,28]. In addition, there is evidence suggesting that this reactivation could lead to the strengthening of the memories (see [2] for a detailed review of the underlying mechanisms). To sum up, such reactivation happen mainly during up-states of SWS, i.e. when most of the neurons are firing in synchrony. In addition, specific high frequency hippocampal oscillations called sharp-wave ripples occur during these up- states, correlate with reactivation events and may induce local LTP. Furthermore, up-states might promote the coordinated action of hippocampal and neocortical networks, allowing transfer of information from one brain region to the other and thus, the long-term storage and stabilization of memories. Finally, REM sleep is thought to be involved to a large extent in the consolidation processes and could favor connections rearrangements [29].
Daily forgetting of irrelevant information and sleep-related selective memory processing are likely to share some characteristics. Activation of relevant neuron patterns occurs in both process and may promote local synaptic strengthening. Altogether, these results lead us to think that SWS might enhance the natural unequal consolidation of memories which were purposely encoded or preferentially retrieved compared to competitors. Therefore, sleep may promote forgetting by resolving interference and reduce the likelihood for irrelevant information to be recalled later. The discrimination process is likely to involve neural circuits in both the hippocampus and the prefrontal cortex. We now have to discuss such selection criteria.
What may determine the information to remember?
We have hypothesized that the relevance of memories may be a substantial element for their over- night consolidation. We will now briefly review to what extent cognitive processes could be involved in this selection. The first point to examine is the subjective relevance that the subject assigns to memories. Given the theoretical sleep-related selective memory consolidation we described, the higher one considers the future importance of some information, the more the memorization of this information is likely to be enhanced by sleep. Two recent studies have been carried out to investigate this hypothesis. Wilhelm et al. [30] found that the retrieval of word-pairs was increased after a sleep period only if the participants were informed of the subsequent recall test after learning. In contrast, Tucker et al. [31] showed that monetary rewarded information (i.e. participants were payed for successful recall) were equally retrieved after a night of sleep than the unrewarded one. Although the subjective will to remember is apparently the same in both Wilhelm and Tucker's studies, their opposite results may be due to the different methodology used (e.g. experimental procedure, items learned...).
On the other hand, the emotional charge of an event might be a significant cue for later memorization. Thus, sleep would especially enhance emotional compared to neutral memories. There is now many evidence that support this hypothesis (see Walker [32] for his works about the role of sleep in emotional memory processing). However, these evidence suggest a specific role of REM sleep rather than SWS in this emotional memory processing. In addition, Walker mentions a hypothetical role of REM sleep in weakening the affective charge of emotional memories (while consolidating the declarative content of these memories). That is, the emotional tone associated with the original emotional event to be remembered would be gradually erased and forgotten, while the factual core of the same event would be strengthened night after night.
A model of sleep-related forgetting
Up until now, we have gathered clues suggesting that sleep could promote a global by default forgetting due to the biochemical and electrical features of SWS. However, all memories would not be affected by such forgetting because of selective consolidation processes occurring during this sleep stage. Although there is large evidence supporting a better strengthening for emotional compared to neutral memories, the other relevance criteria are currently discussed. This form of sleep-related forgetting may be tied to forgetting by resolving interference that occurs every day (i.e. irrelevant information is selected against). The figure 1 strives to sum up the implications of such a sleep-related forgetting model (Figure 1).
Before we discuss its relevance, we have to examine whether behavioral studies fit with this model. While sleep and forgetting have been two distinct fields of research for a long time, the past two years have seen a sudden interest in the role of sleep in forgetting.
Figure 1. A model for a theoretical forgetting function of sleep.
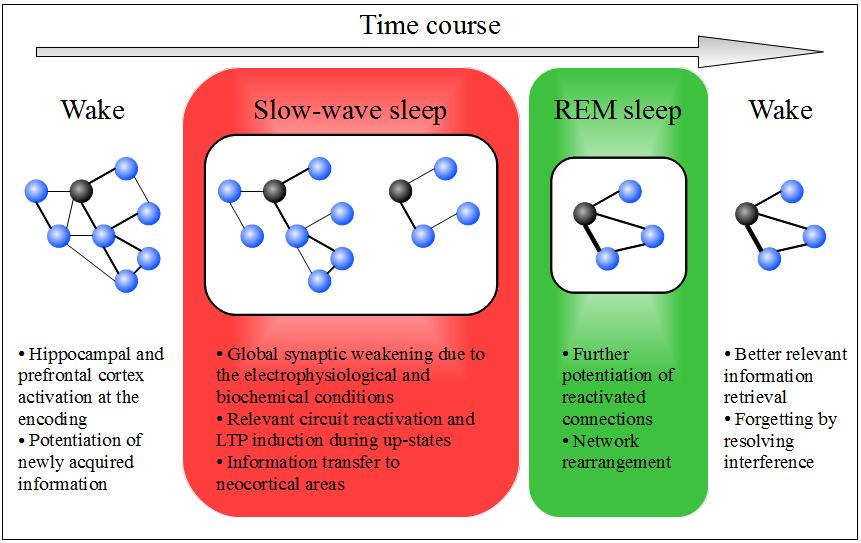
This diagram describes the processing of a relevant memory (symbolized by a black ball, i.e. an interesting paper found among a vast literature) according to the hypothesis that sleep may promote a selective strengthening of this latter and a by default forgetting of competitive memories (symbolized by blue balls). The bounds between the balls represent the associative strength of memories. It is to note that the overnight outcome is a decrease in competitive memories number (the sleep-related forgetting we assess) and an increase in associative strength of the resulting network. However, that should nevertheless lead to a better retrieval of the initially relevant information given that closely related memories are likely to be recall together.
Recent behavioral studies analysis
Several independent approaches but divergent results
One of the first studies which experimentally focuses on both forgetting and sleep was carried out two years ago in human by Racsmány et al. [33] using a retrieval-induced forgetting (RIF) paradigm. In this experiment, participants were instructed to learn associated word-pairs for a subsequent retrieval test. Typically, the first word of the pair (the cue word, e.g. MUSIC) was associated with a semantically-related word (the target word, e.g. violin). Several target words with different first letter were assigned to a same cue word resulting in many groups of pairs : for instance the pairs [MUSIC-violin ; MUSIC-symphony ; MUSIC-rhythm ; MUSIC-drums ; MUSIC- blues ; MUSIC-orchestra] constituted one group. The experiment was divided into three phases: the encoding phase, the practice phase which followed on immediately, and the recall test which was delayed. During the practice phase, some of the groups were tested (i.e. the subject was given a cue word and the first letter of an associated target word and had to recall it (e.g. MUSIC-v for MUSIC- violin) others were not (nRp words; e.g. FRUIT-apple). For each practiced group, some target words were practiced (Rp+ words; e.g. MUSIC-violin), others were not (Rp- words; e.g. MUSIC- drums). Such procedure induces a RIF effect, that is the recall of the Rp+ words interferes with the memorization of the Rp- words, that eventually would cause a worst recall of these words compared to nRp words during the delayed test [34]. In order to assess the effect of sleep on that retrieval- induced forgetting, Racsmány et al. allowed half of the participants to sleep between the practice phase and the recall test whereas the other half stayed awake. They finally reported that RIF was greater in the sleep-group (Figure 2). However, in response to this study, Baran et al. [35] carried out a similar work but found opposite results. They used the same paradigm with more word-pairs and provided participants with feedback during the practice phase (i.e. the subject was informed if he was right or wrong and the target word was given if need was). They concluded with that procedure that RIF decreased after a period of sleep (Figure 2). Moreover, they reported that the percent of the night spent in REM sleep was negatively correlated to the competitive forgetting, that is, REM sleep might reduce forgetting. These two studies have raised controversial issues on the hypothetical forgetting function of sleep that we will discuss below in regards to the works which have followed them.
Figure 2. Controversial results concerning the effect of sleep on retrieval-induced forgetting.
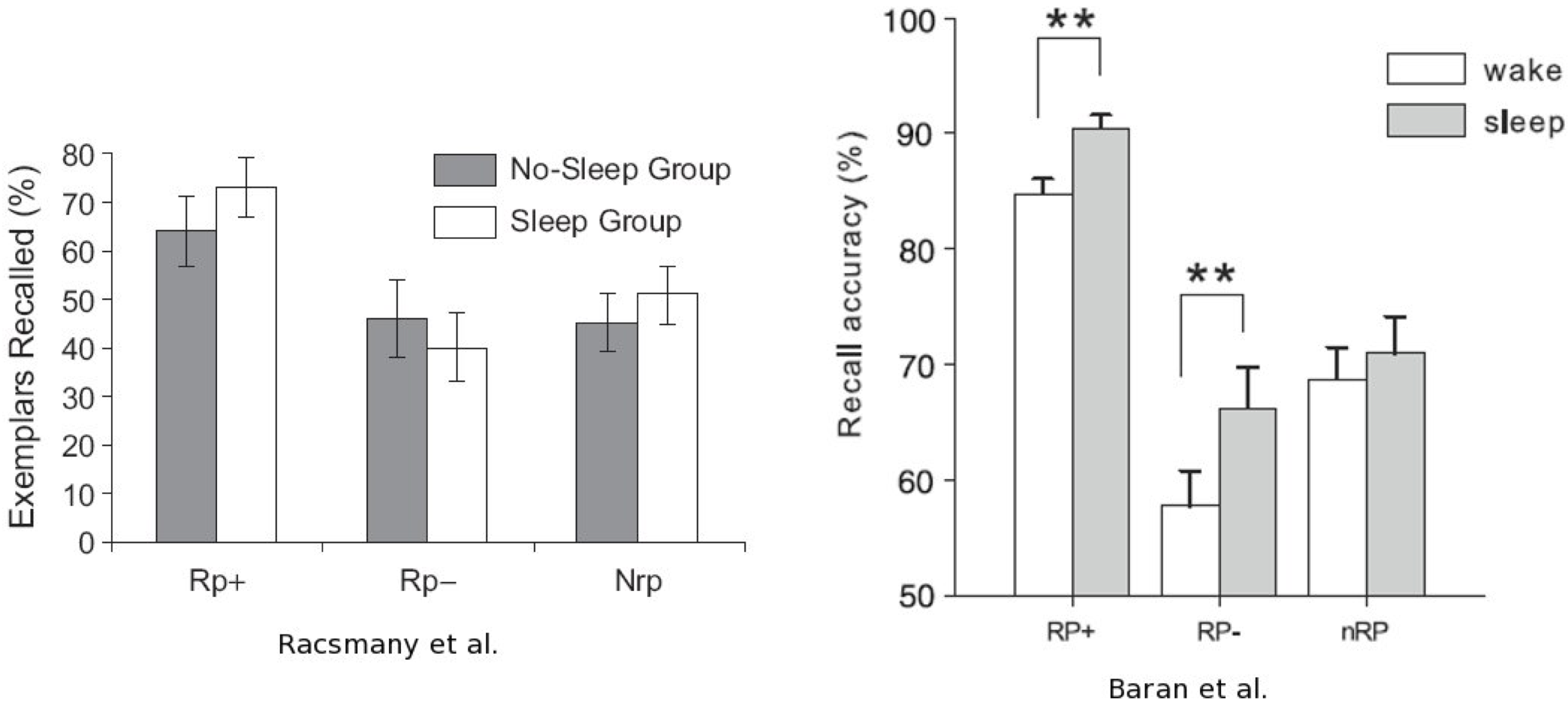
The same RIF paradigm was used in two studies to investigate the effect of sleep on forgetting. Rp+ practiced items ; Rp- unpracticed and selected against items ; nRp unpracticed only items (baseline). The higher is the recall of Rp+ compared to Rp-, the higher is the RIF effect. Racsmány et al. [33] reported a significant RIF effect in the sleep group only (comparison between columns 1 and 3 versus columns 2 and 4). In contrast, Baran et al. [35] did not observe any difference in the RIF level after a period of sleep or wakefulness. Moreover, they noticed that sleep promoted the consolidation of both relevant (Rp+) and competitive (Rp-) memories. (Reproduced from Racsmány et al. [33], 2010 and Baran et al., 2010) [35].
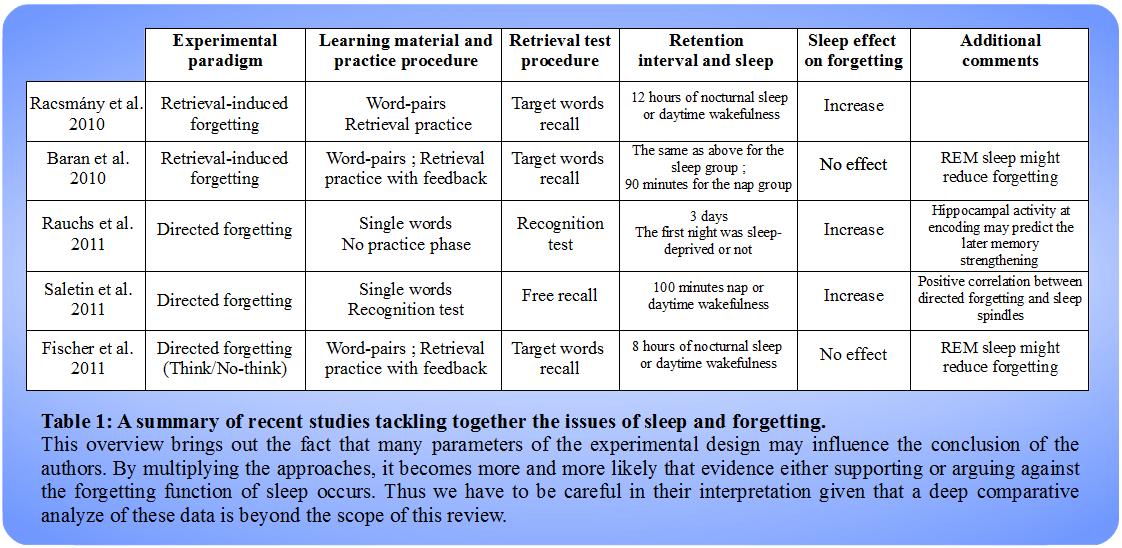
Other studies have tackled the issue differently by using another experimental paradigm called directed forgetting. In this paradigm, the encoding phase consists in succeeding words which are cued to be remembered (TBR words) or to be forgotten (TBF words) (see the works of Kuhl & Wagner [25] for a detailed presentation of the different kinds of forgetting and the associated experimental paradigms). Directed forgetting paradigm relies on the active inhibition of memorization for some items during the encoding phase whereas all the items were supposed to be remembered in the previous retrieval-induced forgetting experiment. Rauchs et al. [36] used this paradigm following by a recognition retrieval task during which participants were asked to tell if they had already seen the words (i.e. during the encoding phase) showed by the experimenter. One group was sleep-deprived the following night. They reported that the sleep-deprived group recognized more TBF words than the regular sleep group whereas the retrieval of TBR words was equal in both the groups suggesting once again that sleep could enhance forgetting of some memories over others. In addition, by using functional magnetic resonance imaging they demonstrated (a) that the encoding of TBR words compared to TBF words correlated with a higher hippocampal activity and (b) that in group that sleep normally, the higher the hippocampal activity at encoding, the better the subsequent retrieval of the item. Therefore, these observations suite well with the hypothesis that sleep might preferentially promote the strengthening of relevant information. Similar results were obtained by Saletin et al. [37] who assigned a free recall test to participants after they had the possibility to have a hundred minutes nap. In this study, the directed forgetting effect was greater in the nap-group despite the fact that the recall of TBF words was equal whether the subjects had slept or not. The free recall of TBR words was indeed enhanced by sleep whereas Rauchs et al. did not find any significant difference about the remembering of TBR words by using their recognition retrieval task. Thus, these two studies came to the same conclusion that sleep could enhance directed forgetting although their results diverged to some extent. In addition, Saletin et al. reported a correlation between the directed forgetting measure and the occurrence of spindles (i.e. local field activity that occurs during the up-states of SWS and might be involved in memory potentiation such as ripples). Moreover, the field was investigated in parallel by Fischer et al. [38] using a variant of the directed forgetting task called the think/no-think paradigm. In this one, to be remembered word-pairs are shown to the participants during a classical encoding phase. Then follows the think/no-think phase during which single words of previously studied pairs are presented to the subjects who alternately have to remember the associated word (i.e. think trial) or to mentally avoid the retrieval of such associated word (i.e. no-think trial). The authors also provided feedback during this phase. They reported that the directed forgetting effect evaluated eight hours later through a retrieval test did not differ from the participants who slept during this period to those who did not. In addition, they found that subjects recalled more TBF items (i.e. no-think word-pairs) after a REM-rich sleep suggesting like the work of Baran et al. that REM sleep could counteracts the voluntary forgetting of unwanted memories.
A synthesis of these studies is offered in Table 1. What stands out is the lack of consensus concerning the possibility for sleep to enhance the forgetting of irrelevant (or to be forgotten) information. This might be due to the variations in experimental procedures whether it involves memory (e.g. forgetting task, practice phase, retention interval, retrieval test) or sleep (e.g. sleep quality, time of sleep). Moreover, human studies always rely to some extent on a subjective approach that could bias the results. For instance, it is difficult to control the individual motivation during the encoding, the rehearsal of information after learning or the sleep quality.
Conclusions
Through this work we have inquired whether the abundant literature relating to sleep and memory is reconcilable with a far less investigated sleep-related forgetting function theory. By gathering evidence from the biochemical, electrophysiological and behavioral studies we hypothesized that sleep may promote forgetting of some memories over others. The model assumes that LTD and synaptic downscaling would by default weaken memories during SWS. On other hand, selective consolidation processes would target memories that are likely to be reactivated during SWS thanks to particular relevance during wakefulness. Thus sleep would enhance a natural mechanism of forgetting by resolving interference. Nevertheless, what determines the relevance of a piece of information remains unclear and behavioral studies failed to provide consistent evidence to support or not this model.
Given the studies analyzed and the restricted scope of this work, it is to note that such a model would account for specific features of both memory and sleep. Indeed, this work mainly refers to declarative hippocampal-dependent memory but procedural memory might undergo different processing. In addition, the role of REM sleep in forgetting has been poorly studied although behavioral studies have suggested that REM sleep might prevent the forgetting of any information, independently of its relevance [35,38]. Therefore, further investigation is needed to better understand what REM-associated processes could explain that overall improvement and to what extent it depends on prior SWS mechanisms.
To conclude, although the increasing interest in the relationship between sleep and forgetting is encouraging, the standardization of experimental procedures (above all in behavioral studies) might be a crucial point in order to challenge the model we have offered and improve our knowledge in both the fields.
Acknowledgments
I would like to thank Philippe Peigneux for having generously given to me his manuscript [3], as well as Paul Salin and Gaël Malleret for their availability and their worthwhile comments.
References and recommended reading
Papers of particular interest have been highlighted as:
● of special interest
●● of outstanding interest
2.Diekelmann S, Born J:
The memory function of sleep.
Nat Rev Neurosci
2010,
11
:114–126.
● An extensive review of the current sleep-related memory function theory.
3.Peigneux P, Schmitz R, Urbain C: Sleep and forgetting. In Forgetting. Edited by Sala S. Psychology Press ; 2010.
4.Levy BJ, Kuhl BA, Wagner AD:
The functional neuroimaging of forgetting.
In
Forgetting.
Edited by Sala S. Psychology Press;
2010.
● The authors reviewed the main mechanisms of forgetting and offered a
classification of its different forms.
5.Kandel ER: The Molecular Biology of Memory Storage: A Dialogue Between Genes and Synapses. Science 2001, 294 :1030–1038.
6.Malinow R, Malenka RC: AMPA receptor trafficking and synaptic plasticity. Annu Rev Neurosci 2002, 25 :103–126.
7.Leonard B, McNaugthon B, Barnes C: Suppression of hippocampal synaptic plasticity during slow-wave sleep. Brain Res 1987, 425 :174–177.
8.Bramham C, Srebro B: Synaptic plasticity in the hippocampus is modulated by behavioral state. Brain Res 1989, 493 :74–86.
9.Pace-Schott EF, Hobson JA: The Neurobiology of Sleep: Genetics, cellular physiology and subcortical networks. Nat Rev Neurosci 2002, 3 :591–605.
10.Vyazovskiy VV, Cirelli C, Pfister-Genskow M, Faraguna U, Tononi G: Molecular and electrophysiological evidence for net synaptic potentiation in wake and depression in sleep. Nat Neurosci 2008, 11 :200–208.
11.Froc DJ, Chapman CA, Trepel C, Racine RJ: Long-Term Depression and Depotentiation in the Sensorimotor Cortex of the Freely Moving Rat. J Neurosci 2000, 20 :438–445.
12.Lanté F, Toledo-Salas J-C, Ondrejcak T, Rowan MJ, Ulrich D: Removal of Synaptic Ca2+- Permeable AMPA Receptors during Sleep. J Neurosci 2011, 31 :3953 –3961.
13.Cirelli C, Gutierrez CM, Tononi G: Extensive and Divergent Effects of Sleep and Wakefulness on Brain Gene Expression. Neuron 2004, 41 :35–43.
14.Ribeiro S, Mello CV, Velho T, Gardner TJ, Jarvis ED, Pavlides C: Induction of Hippocampal Long-Term Potentiation during Waking Leads to Increased Extrahippocampal zif-268 Expression during Ensuing Rapid-Eye-Movement Sleep. J Neurosci 2002, 22 :10914–10923.
16.Nicholls RE, Alarcon JM, Malleret G, Carroll RC, Grody M, Vronskaya S, Kandel ER: Transgenic Mice Lacking NMDAR-Dependent LTD Exhibit Deficits in Behavioral Flexibility. Neuron 2008, 58 :104–117.
17.Malleret G, Alarcon JM, Martel G,
Takizawa S, Vronskaya S, Yin D, Chen IZ, Kandel ER, Shumyatsky GP:
Bidirectional Regulation of Hippocampal Long-Term Synaptic Plasticity and Its
Influence on Opposing Forms of Memory.
J Neurosci
2010,
30
:3813 –3825.
● By using mutant mice the authors investigated the role of LTP and LTD in
different forms of memory. They suggested that LTD may be involved in forgetting
processes and thus allow new information acquisition and behavioral flexibility.
19.Appelbaum L, Wang G, Yokogawa T, Skariah GM, Smith SJ, Mourrain P, Mignot E: Circadian and Homeostatic Regulation of Structural Synaptic Plasticity in Hypocretin Neurons. Neuron 2010, 68 :87–98.
20.Bushey D, Tononi G, Cirelli C:
Sleep and Synaptic Homeostasis: Structural Evidence in
Drosophila.
Science
2011,
332
:1576 –1581.
● The increase in sleep need and synaptic strenght after wakefulness has
been reported in
Drosophila
. Sleep induced synaptic
renormalization which may involve the gene fmr1. These results supported
synaptic homeostasis theory.
21.Borbély AA, Achermann P: Sleep homeostasis and models of sleep regulation. J Biol Rhythms 1999, 14 :557–568.
22.Finelli L., Baumann H, Borbély A., Achermann P: Dual electroencephalogram markers of human sleep homeostasis: correlation between theta activity in waking and slow-wave activity in sleep. Neuroscience 2000, 101 :523–529.
23.Mander BA, Santhanam S, Saletin JM, Walker MP: Wake deterioration and sleep restoration of human learning. Curr Biol 2011 , 21 :R183–R184.
24.Drosopoulos S, Schulze C, Fischer S, Born J: Sleep's function in the spontaneous recovery and consolidation of memories. J Exp Psychol Gen 2007, 136 :169–183.
25.Kuhl B, Wagner A: Forgetting and retrieval. In Handbook of Neurosciences for the Behavioral Sciences. Edited by Berntson G, Cacioppo J. John Wiley and Sons; 2009.
26.Kuhl BA, Shah AT, DuBrow S, Wagner AD: Resistance to forgetting associated with hippocampus-mediated reactivation during new learning. Nat Neurosci 2010, 13 :501–506.
27.Wilson M, McNaughton B: Reactivation of hippocampal ensemble memories during sleep. Science 1994, 265 :676–679.
28.Peigneux P, Laureys S, Fuchs S, Collette F, Perrin F, Reggers J, Phillips C, Degueldre C, Del Fiore G, Aerts J, et al.: Are Spatial Memories Strengthened in the Human Hippocampus during Slow Wave Sleep? Neuron 2004, 44 :535–545.
29.Payne JD: Sleep on it!: stabilizing and transforming memories during sleep. Nat Neurosci 2011, 14 :272–274.
30.Wilhelm I, Diekelmann S, Molzow
I, Ayoub A, Mölle M, Born J:
Sleep Selectively Enhances Memory
Expected to Be of Future Relevance.
J Neurosci
2011,
31
:1563–1569.
●● The authors reported a cross-effect on word-pairs memorization between
sleep and expectancy for a later retrieval. The performance was better in
subjects who both slept and expected the recall test.
31.Tucker MA, Tang SX, Uzoh A, Morgan
A, Stickgold R:
To Sleep, to Strive, or Both: How Best to Optimize
Memory.
PLoS ONE
2011,
6
:e21737.
● In this study, it has been shown that monetary rewarded declarative
memories consolidation does not benefit from sleep.
32.Walker MP, van der Helm E: Overnight Therapy? The Role of Sleep in Emotional Brain Processing. Psychol Bull 2009, 135 :731–748.
33.Racsmány M, Conway MA, Demeter G:
Consolidation of Episodic Memories During Sleep.
Psychol Sci
2010,
21
:80–85.
● This was the first study to give evidence that sleep may promote
forgetting by using a retrieval-induced forgetting paradigm. The
retrieval-induced forgetting in word-pairs recall was higher in subjects who
slept between the learning phase and the recall test.
34.Anderson M, Bjork R, Bjork E: Remembering can cause forgetting - Retrieval dynamics in long-term-memory. J Exp Psychol Learn 1994, 20 :1063–1087.
35.Baran B, Wilson J, Spencer RMC:
REM-dependent repair of competitive memory
suppression.
Exp Brain Res
2010,
203
:471–477.
●● The authors repeated the experiment carried out by Racsmány
et al.
[33] with little differences in the procedure
(more items to learn, feedback during retrieval practice). They reported no
effect of sleep on retrieval-induced forgetting, in opposition to the previous
study. In addition, they found that REM-rich night reduced forgetting.
36.Rauchs G, Feyers D, Landeau B,
Bastin C, Luxen A, Maquet P, Collette F:
Sleep Contributes to the
Strengthening of Some Memories Over Others, Depending on Hippocampal Activity at
Learning.
J Neurosci
2011,
31
:2563 –2568.
●● Sleep effect on forgetting has for the first time been studied using a
directed forgetting paradigm. The recall for the to be forgotten information was
lower in subjects who slept between learning and retrieval phases and correlated
with a lower hippocampal activity at encoding. These results supported the fact
that sleep may promote the forgetting of irrelevant information.
37.Saletin JM, Goldstein AN,
Walker MP:
The Role of Sleep in Directed Forgetting and
Remembering of Human Memories.
Cereb Cortex
2011,
21
:2534 –2541.
●● The authors reported that sleep may enhance directed forgetting of
single words learned before a day nap. Moreover, they found a positive
correlation between directed forgetting level and sleep spindles occurrence in
slow-wave sleep suggesting its importance for memories discrimination.
38.Fischer S, Diekelmann S, Born
J:
Sleep's role in the processing of unwanted memories.
J Sleep Res
2011,
20
:267–274.
●● Using the think/no-think paradigm the authors showed that sleep globally
had no effect on directed forgetting. A REM-rich night may however reduce the
forgetting of to be forgotten memories.